


* Department of Genetics, Washington University School of Medicine, St.
Louis, MO 63110-1095
**Museum of the Mendelianum, Brno, Czechoslovakia
Copyright © 1992 by the Genetics Society of America.
(This paper originally appeared in Genetics 131: 245-253 (1992). It is made available at MendelWeb, for non-commercial educational use only, with the kind permission of Dr. Hartl and Genetics. Although you are welcome to download this text, please do not reproduce it without the permission of the authors and Genetics.)
GREGOR MENDEL is accorded a special place in the history of genetics. His experiments, beautifully designed, were the first to focus on the numerical relationships among traits appearing in the progeny of hybrids; and his interpretation, clear and concise, was based on material hereditary elements that undergo segregation and independent assortment. Poignantly overshadowing the creative brilliance of Mendel's work is the fact that it was virtually ignored for 34 years. Only after the dramatic rediscovery in 1900--16 years after Mendel's death--was Mendel rightfully recognized as the founder of genetics.
Mendelian mythmaking? This orthodox interpretation of Mendel's contribution has recently been challenged as
...a myth created by the early geneticists to reinforce the belief that the laws of inheritance are obvious to anyone who looks closely enough at the problem (Bowler 1989, p. 103).
This opinion reflects revisionist views of Mendel and his intentions that have received considerable prominence in journals dealing with the history of science. The new perspectives come from several directions. One influential view is that Mendel was not interested in heredity, as such, but in the role of hybrids in the generation of new species. This view of Mendel's work
...strips it of inflated whiggish interpretations and places it squarely within the context of mid-nineteenth century biology (Olby 1979, p. 53).
The iconoclastic conclusion is that if you define a Mendelian
...as one who subscribes explicitly to the existence of a finite number of hereditary elements which in the simplest case is two per hereditary trait, only one of which may enter a germ cell, then Mendel was clearly no Mendelian (Olby 1979, p. 68).
A second view echoes the emphasis on Mendel as hybridist
...concerned with the formation and development of hybrids ... using empirical methods ... [who did] not explain his results by employing invisible particulate determiners, paired or otherwise (Monaghan and Corcos 1990, p. 268).
A third view imputes to Mendel a dark motive in performing his experiments: that he had actually set out to prove that hybrids gave invariant progeny (essentially an antidarwinian rejection of descent with modification) and that his work with Hieracium was not undertaken to demonstrate agreement with his results with Pisum but rather to demonstrate the existence of constant (nonsegregating) hybrids (Callender 1988). A fourth view goes well beyond the others in claiming that
[Mendel's monohybrid] experiments are fictitious in the sense that they have been carried out only on paper: the numerical data relative to them have been obtained by progressively disaggregating those from polyhybrid crosses. In other words, Mendel never carried out these experiments in the garden, but rather only on the pages of his notebooks (Di Trocchio 1991, p. 515).
We believe that it would be worthwhile to examine these claims in a forum accessible to geneticists. Most instructors in genetics, and most genetics textbooks, still put forth the orthodox interpretation of Mendel's work. Stern and Sherwood ( 1966), in the foreword to their translation of Mendel's Versuche über Pflanzen-Hybriden, point out that
...in the history of Mendelism the eternal is strongly confounded with the ephemeral. There is the neglect of Mendel's accomplishments during his lifetime and the futility of his years of writing to Nägeli. There is the uneven history of the rediscovery of his work thirty-four years after its publication and its striking reanalysis [by Fisher (1936)] another thirty-six years later.
Only time will tell whether the revisionist views of Mendel's work are ephemeral, but if the orthodox interpretation of Mendel's work is seriously flawed in any respect, then it ought to be adjusted.
Why the neglect? Although the main themes of this paper are paraphrases of Fisher's (1936) searching questions--What was Mendel trying to discover? What did he discover? What did he think he discovered?--there are additional issues concerning Mendel's work that warrant updating. First, why was the work neglected for so long? A comprehensive and thoughtful discussion of various categories of explanation is presented by Sandler and Sandler (1985). While granting a role to the relative inaccessibility of the Verhandlungen des naturforschenden Vereines in Brünn (Brno), in which Mendel's paper was published, to the overshadowing contemporary interest in organic evolution, and to the strangeness and novelty of Mendel's experimental approach, Sandler and Sandler nevertheless conclude that Mendel's paper was simply incomprehensible to his contemporaries because heredity and development were completely confounded conceptually in the latter half of the nineteenth century. The transmission of hereditary traits from parents to offspring was considered part of the same process as the development of traits in the offspring; heredity was but a moment in development with no need for a conceptual framework of its own. Hence, while
Mendel ... defined his problem in purely genetic terms, and produced a correct and amazingly complete answer, [it was] to an as yet unformulated question! (Sandler and Sandler 1985, p. 69).
The all too goodness of fit: Another issue is the excessive goodness of fit of Mendel's ratios (Fisher 1936; Edwards 1986). It is unfortunate that Fisher's
...painstaking analysis and his defense of Mendel's integrity have sometimes been incorrectly reported as having exposed a scientific fraud of major proportions .... (Edwards 1986, p. 296).
The truth is that, while the data show a persistent lack of extreme segregations, as if there had been some trimming of highly deviant values (Edwards 1986), the bias toward expectation is slight (Wright 1966). Considering the entirety of the evidence, Edwards (1986) agrees with Dobzhansky's conclusion:
Few experimenters are lucky enough to have no mistakes or accidents happen in any of their experiments, and it is only common sense to have such failures discarded. The evident danger is ascribing to mistakes and expunging from the record perfectly authentic experimental results which do not fit one's expectations. Not having been familiar with chi-squares and other statistical tests, Mendel may have, in perfect conscience, thrown out some crosses which he suspected to involve pollen contamination or other accident (Dobzhansky 1967, p. 1589).
More recently, Mendel has also been accused of a more subtle kind of deception (Di Trocchio 1991). From his analysis of the Versuche, Di Trocchio supposes that Mendel did not actually do the crosses he reported. What he actually did, according to this reconstruction, was to carry out no fewer than 484 crosses representing all pairwise reciprocal combinations of the 22 varieties of Pisum that Mendel had available. Most of these crosses would have shown either no segregation or segregation for several traits. Among the latter, Mendel pooled the marginal totals and reported them as monohybrid segregations. Even if Mendel followed this procedure, the marginal totals were obtained in real crosses and could hardly be regarded as "fictitious." Furthermore, it is hard to believe that Mendel actually followed this procedure without saying so, since he roundly criticized Gärtner for not describing his own experiments in sufficient detail to allow Mendel to repeat them:
The results which Gärtner obtained in his experiments are known to me; I have repeated his work and have reexamined it carefully to find, if possible, an agreement with those laws of development which I found to be true for my experimental plant. However, try as I would, I was unable to follow his experiments completely, not in a single case! It is very regrettable that this worthy man did not publish a detailed description of his individual experiments....However, in most cases, it can at least be recognized that the possibility of an agreement with Pisum is not excluded (letter to Nägeli, 31 December 1866, p. 57).
Mendel as a hybridist: In considering Mendel's contributions, the greatest weight must be placed on his own writing on hybridization, which is unfortunately limited to two papers (1866 and 1870) and ten letters to Carl Nägeli (1866-1873). The principal source of information is the Versuche (1866), which was the text of Mendel's public lectures of February 8 and March 8, 1865, to fellow members of the Brno Natural History Society. In Mendel's letter of April 18, 1867, to Nägeli, he says,
I made every effort to verify, with other plants, the result obtained with Pisum....I attempted to inspire some control experiments, and for that reason discussed the Pisum experiments at the meeting of the local society of naturalists. I encountered, as was to be expected, divided opinion....When, last year [1866] I was asked to publish my lecture in the proceedings of the society, I agreed to do so, after having re-examined my records for the various years of experimentation, and not having been able to find a source of error. The paper which was submitted to you is the unchanged reprint of the draft of the lecture mentioned; thus the brevity of the exposition, as is essential for a public lecture (p. 60).
Therefore, while the Versuche is written in remarkably clear German (Stern and Sherwood 1966), the account is abbreviated and addressed to a general audience. We have no way of knowing what revisions Mendel might have made had he foreseen that every word and phrase in the paper would still be parsed and analyzed more than 125 years afterward. For example, Di Trocchio (1991, p. 516) readily admits that his allegation that Mendel's monohybrid experiments were "fictitious" is based almost exclusively on the occurrence of two "unexpected" words: eintheilung ("breaking up into several parts"), which appears in the third subheading of the Versuche (p. 5); and wesentlich ("essential, substantial"), which appears in the statement that "plants were used which differed in only one essential trait" (p. 17).
Did Mendel have an agenda to disprove descent with modification that led to his choice of Hieracium, with its constant hybrids, for his later experiments (Callender 1988)? The Darwinian theory was well known by the time of Mendel's presentation. The German translation of On the Origin of Species was published in 1863, and the monastery copy has marginal notes in Mendel's handwriting (1). In January, 1865, at the monthly meeting of the Brno Natural History Society, A. Makowsky, one of Mendel's friends, lectured on the theory of natural selection. Mendel gave the next two lectures, and thus he could easily have focused on the evolutionary angle had he wished. However, the Versuche mentions evolution on only three occasions: on p. 2 in the phrase "the evolutionary history of organic forms," on p. 41 in the phrase "the evolutionary history of plants," and on p. 47 where Mendel cites Gärtner's view that "a species has fixed limits beyond which it cannot change" and says that this statement cannot be accepted unconditionally (2). Furthermore, a number of plant species other than Hieracium were chosen in making "every effort to verify, with other plants, the result obtained with Pisum" (Mendel's letter to Nägeli 18 April 1867, p. 60). The reason Mendel chose Hieracium is given in his 1870 paper:
This genus possesses such an extraordinary profusion of distinct forms that no other genus of plants can compare with it (p. 51).
Mendel's overriding interest was
...the question whether and to what extent hybridization plays a part in the production of this wealth of forms (p. 51)
knowing full well that
...we may be led into erroneous conclusions if we take rules deduced from observation of certain other hybrids to be laws of hybridization, and try to apply them to Heiracium without further consideration (p. 52).
At the beginning of his Pisum experiments Mendel was certainly aware of the Linnean concept of constant hybrids and thought that they might be important exceptions to the laws deduced for Pisum. In the Versuche he remarked that there is
... an essential difference in those hybrids that remain constant in their progeny and propagate like pure strains (p. 41, emphasis his)
He also understood that
...this feature is of particular importance to the evolutionary history of plants, because constant hybrids attain the status of new species (p. 41, emphasis his).
On the other hand, he had
...proved experimentally that in Pisum hybrids form different kinds of germinal and pollen cells and that this is the reason for the variability of their offspring (p. 41, emphasis his).
Far from providing the constant hybrids that Mendel was allegedly seeking, the experiments with Hieracium, as recounted in the letters to Nägeli, were one long chronicle of failure and frustration.
On the other hand, there is no disputing that Mendel was in the tradition of the plant hybridists, like Kölreuter and Gärtner, whose work, among that of others, he cites in the Versuche. The question is whether he was so engrossed in this tradition that the process of heredity itself was either unrecognized or of little interest (Olby 1979, Monaghan and Corcos 1990). Olby (1979, p. 67) claims that
Mendel's overriding concern was with the role of hybrids in the genesis of new species....The laws of inheritance were only of concern to him in so far as they bore on his analysis of the evolutionary role of hybrids.
It is certainly true that Mendel's main interest was in hybrids. The answer to the question "What was Mendel trying to discover?" is clearly given in the introductory remarks in the Versuche, in which Mendel describes the experimental program necessary to discover a
...generally applicable law of the formation and development of hybrids (p. 2)
which he considered as a law whose
...significance for the evolutionary history of organic forms must not be underestimated (p. 2).
On the other hand, in the very first sentence of the Versuche, Mendel explains that the experiments were based on
...artificial fertilization undertaken on ornamental plants to obtain new color variants (p. 1),
which seems also to imply a significant interest in heredity for its role in practical breeding. Mendel also had little concern for the systematic status of his pea plants. He remarks that, in the opinion of experts, most of the plants belong to the species Pisum sativum, but
...the rank assigned to them in a classification system is completely immaterial to the experiments in question (p. 5).
One will never know exactly what Mendel was thinking, but one might reasonably have expected a less cavalier attitude toward species classification on the part of an author whose primary concern was the role of hybridization in the formation of new species. (Mendel was not alone in regarding the distinction between species and varieties as purely arbitrary. The view was common among 19th century biologists, including Darwin, and almost universal among the botanists. It was to cause Mendel great trouble after 1866 when he began making hybrids between what would now be regarded as distinct species. We are indebted to Ernst Mayr for pointing this out in a letter of 17 December 1991 to DLH.)
The Moravian connection: If, as Sandler and Sandler (1985) point out, there was no clear conceptual distinction between heredity and development, then how could Mendel have been interested in heredity for its own sake? Even before Mendel's time, there had been great interest in Moravia in the practical breeding of sheep, fruit trees, and vines, partly through the leadership of the naturalist, C. C. André (1763-1831), who in 1806, through the patronage of Count Salm-Reifferscheid (1776-1836), had organized in Brno the Royal and Imperial Moravian and Silesian Society for the Improvement of Agriculture, Natural Science and Knowledge of the Country (Orel 1983). Many of the monks of the monastery of St. Thomas were active in this society, including Mendel's predecessor as abbot, F. C. Napp (1792-1867), and Mendel himself. Napp had organized the Brno Natural History Society as one of the sections in the larger organization until it become independent in 1861 (3). If Mendel had a mentor in Brno, it was Napp: he had supported the teaching of agriculture in the context of natural science, and Mendel attended these lectures in 1846; and he had Mendel sent to the university in Vienna in 1851-1853 to study with, among others, the prominent plant physiologist F. Unger and the physicist C. J. Doppler (of the Doppler effect) (4). On the other hand, Napp's role in shaping Mendel's outlook and research agenda is uncertain. What is clear is that Napp, motivated by issues in practical breeding, was interested in heredity as a problem in itself. In 1836 he attended a meeting of the Sheep Breeders Association and summarized the discussion of breeding methods by noting that the crucial questions were: What is inherited? How is it inherited? (Orel 1984). Later, in 1840, at a meeting of agriculturalists, Napp defended hybridization as a method of obtaining new varieties of fruit trees and drew attention to the element of chance in this process (Orel 1983). That Mendel himself was interested in practical breeding can hardly be doubted, since he chose to memorialize these interests in ceiling paintings on the renovated monastery reception room as well as on his own tomb (Matalová 1983). Mendel even ate the fruits of his pea breeding efforts:
In 1859 I obtained a very fertile descendant with large, tasty, seeds from a first generation hybrid. Since, in the following year, its progeny retained the desirable characteristics and were uniform, the variety was cultivated in our vegetable garden, and many plants were raised every year up to 1865 (Mendel letter of 18 April 1867 to Nägeli, p. 61).
What did Mendel discover? If Mendel was trying to discover a "generally applicable law of the formation and development of hybrids," what did he actually discover? In the traditional view, Mendel discovered that hereditary traits are determined by cellular elements, now called genes, that exist in pairs, undergo segregation and independent assortment, and persist unchanged through successive generations of hereditary transmission. A key passage in the Versuche that is relevant to this interpretation is as follows (p. 29ff):
The difference of forms among the progeny of hybrids, as well as the ratios in which they are observed, find an adequate explanation in the principle [of segregation] just deduced. The simplest case is given by the series for one pair of differing traits. It is shown that this series is described by the expression: A + Aa + a, in which A and a signify the forms with constant differing traits, and Aa the form hybrid for both. The series contains four individuals in three different terms. In their production, pollen and germinal cells of form A and a participate, on the average, equally in fertilization; therefore each form manifests itself twice, since four individuals are produced. Participating in fertilization are thus:
Pollen cells: A + A + a + a
Germinal cells: A + A + a + a
It is entirely a matter of chance which of the two kinds of pollen combines with each single germinal cell. However, according to the laws of probability, in an average of many cases it will always happen that every pollen form A and a will unite equally often with every germinal-cell form A and a; therefore, in fertilization, one of the two pollen cells A will meet a germinal cell A, the other a germinal cell a, and equally, one pollen cell a will become associated with a germinal cell A, and the other a.
Pollen cells A A a a | \ / | | X | | / \ | Germinal cells A A a aThe result of fertilization can be visualized by writing the designations for associated germinal and pollen cells in the form of fractions, pollen cells above the line, germinal cells below. In the case under discussion one obtains
A/A + A/a + a/A + a/a
In the first and fourth terms germinal and pollen cells are alike; therefore the products of their association must be constant, namely A and a; in the second and third, however, a union of the two differing parental traits takes place again, therefore the forms arising from such fertilizations are absolutely identical with the hybrid from which they derive. Thus, repeated hybridization takes place. The striking phenomenon, that hybrids are able to produce, in addition to the two parental types, progeny that resemble themselves is thus explained. A/a and a/A both give the same association Aa, since, as mentioned earlier, it makes no difference to the consequence of fertilization which of the two traits belongs to the pollen and which to the germinal cell. Therefore
A/A + A/a + a/A + a/a = A + 2Aa + a
This represents the average course of self-fertilization of hybrids when two differing traits are associated in them. In individuals flowers and individual plants, however, the ratio in which the members of the series are formed may be subject to not insignificant deviations.
In our opinion, this passage is strikingly perceptive and hardly betrays befuddlement or deep confusion as to whether the formative elements are paired in the plants and segregate in the reproductive cells. Although the passage does not use any of the words gene, genotype, phenotype, homozygous, heterozygous, or segregation, its meaning to modern geneticists is unmistakable. The only fault, which occurs on several occasions in the Versuche, is that the term trait (Merkmal) is used to mean either phenotype or allele, depending on the context, which indicates that the distinction may not always have been clear or entirely sharp in Mendel's own mind.
Particulate inheritance: At this point it is necessary to discuss several issues bearing on the level of Mendel's understanding of his own work (Olby 1979, Meijer 1983, Monaghan and Corcos 1990, Di Trocchio 1991). One issue is whether Mendel thought of his "potentially formative elements (bildungsfähigen Elemente)" (Versuche, p. 43) as particles rather than as fluids or emulsions (Meijer 1983, Monaghan and Corcos 1990). Nowhere in the Versuche is the physical nature of die Elemente discussed in enough detail to infer how Mendel might have imagined them. On p. 42, in the concluding remarks, Mendel refers to "the material composition and arrangement of the elements...in the cell" ("in der materiellen Beschaffenheit und Anordnung der Elemente...in der Zelle"). This phrase would at least seem to imply that he was thinking in terms of some sort of material entities, but even this interpretation is disputed by Kalmus (1983), who dismisses it as "an afterthought" (p. 61) and argues instead that Mendel must have been thinking of die Elemente in terms of nonmaterialistic scholastic metaphysics and invoking the Aristotelian concept of the potential. But this opinion seems flatly contradicted by Mendel's unselfconsciously materialistic statement that
...the distinguishing traits of two plants can, after all, be caused only by differences in the composition and grouping of the elements existing in dynamic interaction in their primordial cells (p. 43)
It does not matter whether Mendel was thinking in terms or particles or fluids, since he emphasized repeatedly the key point that differing elements emerge unchanged from their association. For example, in the letter of April 18, 1867, to Nägeli, Mendel wrote,
The course of development consists simply in this; that in each generation the two parental traits appear, separated and unchanged, and there is nothing to indicate that one of them has either inherited or taken over anything from the other (p. 62).
(This passage also contains Mendel's only known use of the term inherited.)
The use of A instead of AA: Mendel routinely used the symbol A in genetic formulas in which a modern geneticist would use AA, and this notation has been interpreted as meaning that Mendel did not believe that hereditary elements occur in pairs (Olby 1979, Monaghan and Corcos 1990). However, throughout most of the Versuche, Mendel used the symbols A and a in a very different sense than used in modern genetics with reference to genes. In his usage, A refers to a plant that breeds true for the dominant trait, and similarly with a for the true-breeding recessive. Occasionally he used the same symbols to refer to the hereditary determinants, as in the quotation above from p. 29ff, and in the context of this particular discussion it is quite clear that the expression
A/A + A/a + a/A + a/a = A + 2Aa + a
summarizes the expected genetic constitutions of the progeny on the left and gives their physical and breeding characteristics on the right. There are also other interpretations of Mendel's symbolism, including that of Meijer (1983), who argues that Mendel's symbol A means "the potential for creating the dominant trait," and hence the use of AA for the homozygote would be redundant, since the two A 's are logically equivalent. Lest we be too harsh in our anachronous criticism of Mendel's somewhat inconsistent use of symbols, it is worthwhile to bear in mind that modern Drosophila geneticists routinely use unpaired symbols when referring to homozygous recessives; for example, in referring to Drosophila strains, the symbol al means the genotype al/al, and cn bw means the genotype cn bw/cn bw.
Segregation in AA and aa genotypes: The segregation issue provides one of Olby's (1979) principal arguments that Mendel was no Mendelian:
[Mendel] did not conceive of pairs of elements in the cell representing and determining the pairs of contrasted characters. If he had this conception he would have allowed a separation between like members of such pairs as well as between unlike members (Olby 1979, p. 66, emphasis his).
The strongest evidence in the Versuche in support of Olby's view is the following passage:
[The] differing elements [in hybrids] succeed in escaping from the enforced association only at the stage at which the reproductive cells develop. In the formation of these cells all elements present participate in completely free and uniform fashion, and only those that differ separate from each other (p. 43).
On the other hand, the context of this passage clearly refers to hybrids that produce variable progeny. Mendel never specifically addresses the question whether segregation occurs in homozygous genotypes, and we have no way of knowing how he might have responded if queried directly about the matter. Mendel's seeming indifference to segregation in homozygotes was entirely consistent with his primary interest in hybrids and the principle that the hereditary determinants emerge unchanged after their association together in hybrids. As Ernst Mayr has noted, "The homozygotes, not being hybrids, simply did not interest him" (personal communication to DLH 28 October 1991).
Also against Olby's argument is Mendel's formula
A/A + A/a + a/A + a/a = A + 2Aa + a
which clearly implies that the homozygous forms A and a each contain two hereditary determinants. In addition, Mendel writes (p. 41),
...it seems permissible to assume that the germ cells of those [plants] that remain constant are identical, and also like the primordial cell of the hybrid.
The key question is whether the word identical (gleichartig) is intended to mean "identical in number" or "identical in type." We presume that Mendel meant identical in both senses, otherwise some qualification would have been included, which would imply that the germ cells from homozygotes must contain only one of each of the paired determinants, since this is the case in "the primordial cell of the hybrid."
In a wider sense, whether or not segregation may be said to occur in homozygous genotypes is largely a matter of semantics. In the contemporary mind segregation is often confused with the process of chromosome separation during meiosis, but Mendel knew nothing about chromosomes. Furthermore, in a precise technical sense, chromosomes undergo disjunction, not segregation. Segregation is a formal genetic phenomenon in which alleles are separated from one another and distributed into different germ cells (Rieger, Michaelis and Green 1968; King and Stansfield 1985). However, by definition, alleles must be different:
Allele...one of two or more alternate forms of a gene occupying the same locus on a particular chromosome ... and differing from other alleles of that locus.... (Rieger, Michaelis and Green 1968, p. 11)
Allele...one of a series of possible alternative forms of a given gene ..., differing in DNA sequence ...." (King and Stansfield 1985, p. 14).
Hence, Mendel's view of segregation occurring only in the heterozygotes (i. e., with different alleles) could easily be defended as being completely consistent even with the modern use of the term. Moreover, this usage provided Mendel with the opportunity to summarize in the following way what he clearly regarded as his main result:
[Pea] hybrids form germinal and pollen cells that in their composition correspond in equal numbers to all the constant forms resulting from the traits united through fertilization" (p. 29, emphasis his).
This summary is the subject of further discussion below.
The Phaseolus issue: In the Versuche Mendel also commented on his results with flower color in Phaseolus multiflorus, in which the F2 generation from an original cross of white X crimson consisted of a whole range of colors from purple to pale violet and white, in the ratio 30 colored : 1 white, instead of the expected 3 : 1. He then wrote,
But these puzzling phenomena, too, could probably be explained by the law valid for Pisum if one might assume that in Ph. multiflorus the color of flowers and seeds is composed of two or more totally independent colors that behave individually exactly like any other constant trait in the plant. With blossom color A composed of independent traits A1 + A2 + ..., which produce the overall impression of crimson coloration, then, through fertilization with the differing trait of white color a, hybrid associations A1a + A2a + ... would have to be formed... (p. 35).
Hence, considering only A1 and A2, the
...terms of the [F2 progeny] series can enter into 9 different combinations, each of which represents the designation for another color:
1 A1 A2 2 A1a A2 1 A2 a 2 A1 A2a 4 A1a A2a 2 A2a a 1 A1 a 2 A1a a 1 a aThe numbers preceding the individual combinations indicate how many plants of corresponding coloration belong to the series" (p. 36).
Olby (1979) discusses this example in some detail and wonders
...why [Mendel] made no apology for putting both A1 and A2 with the same contrasted character a....The chief reason for this obscurity was ... that Mendel was thinking in terms of the white colour when he wrote down a....It may, of course, be objected that the way Mendel set out his Phaseolus series [the matrix above] shows that he was thinking of two gene loci for white, hence the genotype a a. ... [However, the layout of the table] makes clear the derivation of the classes of offspring from the multiplication of terms and no more (Olby 1979, p. 60ff).
As is clear from Olby's discussion, there is plenty of room for disagreement about Mendel's intentions in summarizing the Phaseolus situation in this manner. Although Mendel undoubtedly associated the symbol a with the color white, he also clearly stated that the two or more colors are "totally independent," that they "behave individually exactly like any other constant trait in the plant," and that "hybrid associations A1a + A2a + ... " would be formed by fertilization with germ cells from a white plant. These comments, and particularly the latter formula, clearly suggest that he had independent factors in mind, and that the a written as a partner for A1 is different from the a written as a partner for A2. Mendel's matrix and the symbol a a makes perfect sense if he were using positional notation, otherwise consistency would require the white plants to be symbolized simply as a.
At the risk of seeming whiggish, we will point out here that positional notation is commonly used in modern Drosophila genetics, although the wildtype alleles, rather than the mutant alleles, are designated with the single symbol, namely +. For example, to a modern Drosophila geneticist, the symbol + + +/y w f clearly means the triple heterozygote for y, w, and f, without any ambiguity, and + w + is the double recombinant chromosome with the wildtype alleles of y and f. We believe that it was this kind of positional notation that Mendel had in mind when he wrote a a in his table of F2 progeny in Phaseolus.
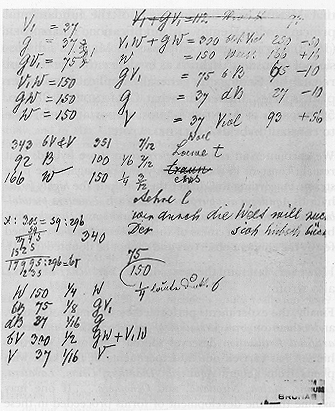 Figure 1:
The Mendel Notizblatt, containing apparent genetic symbols in Mendel's
handwriting, dated to about 1875.
Figure 1:
The Mendel Notizblatt, containing apparent genetic symbols in Mendel's
handwriting, dated to about 1875.
The Notizblatt argument: A good deal of discussion has centered on interpreting what appear to be genetic symbols in Mendel's handwriting found on a document called the Mendel Notizblatt (Figure 1), which bears the rough draft of a letter on the overleaf containing material that dates the letter (though possibly not the putative genetic symbols) to about 1875, or some seven years after Mendel became abbot and two years after his last crossing experiments (Richter 1924). It is not at all clear what the symbols in the Notizblatt refer to: results from Linaria, Phaseolus, and Pisum have all been suggested (Heimans 1968). The Pisum suggestion comes from Heimans (1968) and is based on the observation that the number of individuals in the class W in the Notizblatt (166) agrees with the total for white seed coat color in the trifactorial cross in the Versuche (Mendel, p. 21). The agreement could be coincidental, since the totals do not agree (601 progeny recorded in the Notizblatt versus 639 in the trifactorial cross). Whatever the case, the Notizblatt suggests that Mendel was playing with the data, apparently trying various ways to group the phenotypes into classes, and he flirts briefly with a 7:3:2 ratio in the middle of the page (Figure 1). However, by the bottom of the page he has gotten to the ratio 1/4 : 1/8 : 1/16 : 1/2 : 1/16, which is a perfectly respectable Mendelian ratio found among the 147 theoretically possible ratios in the F2 progeny of a dihybrid cross (Hartl and Maruyama 1968).
It should also be noted that 7:2:3 is quite close to 9:3:4, which is one of the conventional examples of a modified 9:3:3:1. Nevertheless, much has been made of the 7:2:3 ratio, and Olby (1979, p. 62) develops a model of what Mendel might have been thinking when he wrote the markings and explains the model in a paragraph that Olby imagines Mendel might have written were this the model that Mendel actually had in mind. However, Meijer (1983) points out that Olby's model does not really explain the 7:2:3 ratio anyway. These disagreements only serve to highlight the main problem with the Notizblatt: it is an unpublished page of markings pertaining to unknown traits in an unidentified organism written at an undetermined time and apparently of little importance to Mendel himself since he used the page as scrap for drafting a letter about monastery business. Given these uncertainties, the Notizblatt is hardly the sort of thing to rely on in making inferences about Mendel's understanding of his work in Pisum (5).
The issue of constant hybrids: In his argument that Mendel was an opponent of the fundamental principle of descent with modification, Callender (1988) also puts great emphasis on Mendel's discussion of "constant" hybrids as evidence that he did not regard his findings as generally applicable to other plant species. Indeed, in what Callender (1988, p. 56) regards as Mendel's "most important reference to constant hybrids," Mendel wrote:
We encounter an essential difference in those hybrids that remain constant in their progeny and propagate like pure strains. According to Gärtner these include the highly fertile hybrids Aquilegia atropurpurea-canadensis, Lavatera pseudolbia-thuringiaca, Geum urbano-rivale, and some Dianthus hybrids....The correctness of these observations is vouched for by the eminent observers and cannot be doubted (p. 41).
However, later in the text of the Versuche, Mendel also wrote:
Finally, the experiments performed by Kölreuter, Gärtner, and others on transformation of one species into another by artificial fertilization deserves special mention....Gärtner himself has carried out 30 experiments of this kind with plants from genera Aquilegia, Dianthus, Geum, Lavatera, Lychnis, Malva, Nicotiana, and Oenothera.... If one may assume that the development of forms proceeded in these experiments in a manner similar to that for Pisum, then the entire process of transformation would have a rather simple explanation (p. 44).
It is important to emphasize that, in this passage, Mendel pointedly and specifically includes all of the genera referred to earlier as containing constant hybrids as among those genera in which one may assume that the laws for Pisum hold.
While Mendel may have been uncertain about the universal applicability of the laws for Pisum, particularly with regard to species with constant hybrids, he had already convinced himself that they were probably valid at least for all variable hybrids, since he asserted:
Whether variable hybrids of other plant species show complete agreement in behavior also remains to be decided experimentally; one might assume, however, that no basic difference could exist in important matters since unity in the plan of development of organic life is beyond doubt (p. 43, emphasis his).
What did Mendel think he discovered? Whatever Mendel thought he had discovered, he was certainly under the impression that it was important. In his letter of April 18, 1867, to Nägeli he says,
I knew that the results I obtained were not easily compatible with our contemporary scientific knowledge, and that under the circumstances publication of one such isolated experiment was doubly dangerous; dangerous for the experimenter and for the cause he represented (p. 60).
In a paper containing as much data and interpretation as the Versuche, how should one decide what the author regarded as the main message? In our opinion, the key is found in the fact that the Versuche was written for oral presentation, and in an oral presentation the principal technique for emphasis is repetition. Thus, in order to discern what Mendel regarded as the main message, one might examine the Versuche in order to identify any particular ideas or passages that are repeated. We have examined the document with this goal in mind, and have identified one idea that is expressed repeatedly in almost identical language. One version of the idea is as follows:
...pea hybrids form germinal and pollen cells that in their composition correspond in equal numbers to all the constant forms resulting from the combination of traits united through fertilization (p. 29, emphasis Mendel's).
This statement, or close variants of it, is repeated no fewer than six times in the Versuche, on pages 24, 29, 32, twice on page 43, and 44. Furthermore, since Mendel refers to this formulation as the "law of combination of differing traits according to which hybrid development proceeds" (p. 32), we may infer that he considered this statement as summarizing the "generally applicable law of the formation and development of hybrids" (p. 2), the elucidation of which he clearly regarded as his main goal of his experiments. That Mendel regarded this law as widely applicable is supported by his statement that it
...remains more than probable that a factor that so far has received little attention is involved in the variability of cultivated plants....[Our] cultivated plants, with few exceptions, are members of different hybrid series whose development along regular lines is altered and retarded by frequent intraspecific crosses (p. 37ff, emphasis his).
The context of this paragraph clearly points to segregation as "the factor that so far has received little attention."
Mendel's own one-sentence summary of what he thought he discovered is remarkably general and concise. It contains both the laws of segregation and independent assortment in a form that is generally applicable to hybrids of any number of unlinked genes. Even a modern geneticist would be hard pressed to draft a sentence of comparable clarity without using terminology unavailable to Mendel.
Conclusions: Our review of the issues and evidence regarding Mendel as a Mendelian leads us to the following conclusions.
* Mendel should certainly be regarded as a hybridist. He was in the hybridist tradition, and this is apparently how he regarded himself. However, his interest in hybridization did not blind him to the recognition of heredity as a process distinct from evolution. His outlook was conditioned by being in Brno, where there was a particular interest in the results of artificial fertilization [slip of the pen...mean artificial selection] for the improvement of sheep, fruit trees, and vines, in which colleagues of Mendel, such as Abbot Napp, were leading participants, and conditioned also by Mendel's own admitted interest in practical breeding as well as the coloration of ornamental flowers. On the other hand, Mendel's hybridist tradition did result in his failure to emphasize the genetic composition of the constant (homozygous) forms, although the symbolism he used in the Versuche
A/A + A/a + a/A + a/a = A + 2Aa + a
implies to us that he was quite aware that the constant forms must contain paired hereditary determinants.
* Nowhere did Mendel stipulate that the hereditary determinants are particulate, as opposed to fluid, vapor, colloid, gel, plasma, or whatever. He did, however, assume that they are material entities, rather than metaphysical "potentials," and asserted that the distinguishing traits of plants are caused only by differences in the composition and grouping of these hereditary elements.
* Mendel clearly considered segregation as a phenomenon that characterizes the hybrids. On the other hand, the issue whether segregation of genes, as abstract entities, can be said to occur in homozygous genotypes is largely a matter of semantics and can be debated even today. For Mendel, the clear cut segregation in heterozygotes carried the critical implication that the hereditary elements remain unchanged by their association together in heterozygous genotypes.
* The laws of inheritance for Pisum were regarded by Mendel as quite generally applicable. He proposed explicitly that they would apply to all forms of variable (segregating) hybrids, and he also suggested that they might very well explain why genera that include constant (nonsegregating) hybrids can be transformed into new forms by means of artificial fertilization. According to Mendel's own words, his studies of Hieracium and other species were undertaken in order to "verify, with other plants, the result obtained with Pisum" (Mendel's letter to Nägeli of 18 April 1867, p. 60).
* The key point of Mendel's discovery is contained in a sentence that he repeated with minor variations no less than six times in the Versuche:
...pea hybrids form germinal and pollen cells that in their composition correspond in equal numbers to all the constant forms resulting from the combination of traits united through fertilization (p. 29, emphasis his).
This is the kind of "law of the formation and development of hybrids" (p. 2) that he had set out to discover, it is a remarkably concise statement of what he actually did discover, and it certainly represents the distillation of what he thought he discovered. We conclude that Mendel understood very clearly what his experiments meant for heredity. He deserves, not only the eponymous credit for Mendelism, but also the historical credit and a considerable measure of respect and admiration for his remarkable insights.
Notes:
1. The first German translation appeared in 1862. Mendel possessed the second edition from 1863. (Orel)
2. A more appropriate translation of Entwicklungsgeschichte is "developmental history" (Orel).
3. This should be the Natural Science Section inside of the Agricultural Society (Orel).
4 Napp sent Mendel to study exact physics with Professor Doppler. (Orel)
5. We shouldn't dismiss the Notizblatt as "apparently of little importance." Weiling has recently suggested that the notes were written in 1880 or later. (Orel)